
Векторы фитопатогенных рисков
Author(s): проф.д-р Оля Караджова, ИПАЗР “Н.Пушкаров” в София; Марияна Лагинова, директор в ЦЛКР
Date: 21.06.2016
5259

Одной из основных проблем сельского хозяйства в нынешнем веке является обеспечение продовольствием постоянно растущего населения планеты за счет повышения урожайности с использованием устойчивого, экологически безопасного подхода. В 2015 году население мира составляло 7,4 миллиарда человек, но, по прогнозам Всемирной организации здравоохранения (ВОЗ), к 2050 году оно увеличится до 9 миллиардов (ООН, 2007). По данным Джорджа Агриоса (1997), вредители сельскохозяйственного производства (до и после сбора урожая) снижают мировую урожайность примерно на 40%.
В последние годы возникли эпидемии новых и ранее известных болезней растений. Они наносят значительный ущерб на новых территориях, которые они захватывают, в результате изменений вирулентности и агрессивности патогена, спектра растений-хозяев, степени зараженности и т.д. Большое количество этих патогенов высокого риска (вирусы, микоплазмы, бактерии) переносятся насекомыми (тли, цикадки, белокрылки, трипсы), клещами и нематодами. Поэтому точная идентификация патогенов и переносчиков, знание эпидемиологии патогенов, биологии и поведения переносчиков, эффективности передачи патогена и определяющих ее факторов являются ключевыми предпосылками для успешного осуществления мероприятий в рамках систем контроля фитопатогенов.
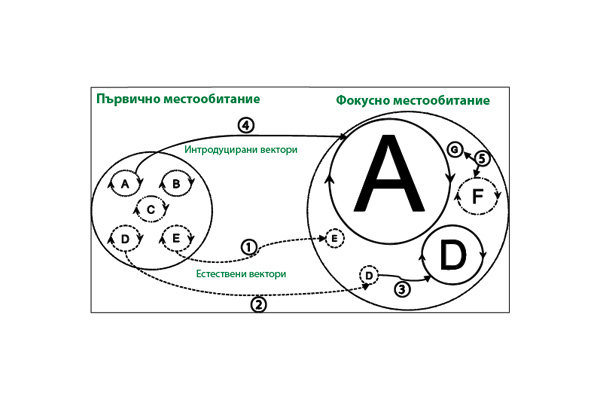
Потенциал возникновения эпидемий заболеваний у людей, животных и растений в результате интродукции переносчиков на новые территории показан на рис. 1.
На левой стороне рисунка представлены штаммы патогенов (A, B, C, D, E), которые поддерживаются в эндемичных циклах болезни в первичных местообитаниях различными типами переносчиков (пунктирные линии в круге). Интродуцированные переносчики представлены сплошными линиями, а местные (аборигенные) – пунктирными. На правой стороне серые круги представляют эпидемические циклы болезни у потенциальных хозяев (люди, животные, растения), причем размер круга соответствует масштабу эпидемии. В этом сценарии в эндемичном цикле несколько патогенных штаммов поддерживаются в первичных хозяевах в течение неопределенного времени местными переносчиками. Иногда местные переносчики случайно заражают конечного хозяина в эпидемическом цикле (1) или распространяются на других хозяев (2). Кроме того, местный переносчик может приобрести патоген от хозяина в эпидемическом цикле и, заразив нового хозяина, вызвать крупномасштабную эпидемию (см. D (3)). Когда интродуцированные переносчики (4) приобретают местный патоген, эпидемия может развиться у нового хозяина (см. A), которая имеет огромные масштабы. Также возможно, что интродуцированные переносчики могут занести совершенно новые патогены (5) в очаговое (местное) местообитание, что может вызвать эпидемии у новых хозяев (F) или заразить хозяев без серьезных последствий (G). Возможна и обратная передача патогенов переносчиками – от эпидемических циклов к эндемическим, создавая резервуары патогенов для будущих эпидемий.
Примером возникновения эпидемии нового заболевания после интродукции нового переносчика является болезнь Пирса винограда в Калифорнии, США (Almeida et al., 2005). Возбудителем болезни является бактерия Xylella fastidiosa, и, хотя патоген присутствует в Калифорнии более 100 лет, за этот период зарегистрировано только три крупные эпидемии. Первая произошла в конце 1800-х годов в Южной Калифорнии и нанесла огромный ущерб виноградникам. Вторая эпидемия болезни Пирса произошла между 1930 и 1940 годами в Центральной Калифорнии и была связана с зараженными цикадками, мигрировавшими с полей люцерны, расположенных рядом с виноградниками. В последующие годы болезнь Пирса винограда обнаруживалась с низкой частотой в прибрежных долинах Напа и Сонома. Третья крупная эпидемия была зарегистрирована в 1999 году, после интродукции в 1989 году в Южную Калифорнию инвазивного переносчика – цикадки Homalodisca vitripennis (Hemiptera: Cicadellidae). В новом местообитании H. vitripennis комплекс естественных врагов отсутствует, и вид размножается до очень высокого уровня популяции. Наличие большого количества переносчиков является основной предпосылкой для эпидемий болезни Пирса винограда в Южной Калифорнии и в самой южной части Центральной долины. Большое количество особей H. vitripennis зимует на цитрусовых (тысячи особей на одно растение). По сравнению с другими видами цикадок, H. vitripennis также характеризуется способностью очень быстро расселяться. Были выявлены два цикла распространения патогена на виноградниках – первый весной, когда взрослые особи мигрируют из цитрусовых садов на виноградники, и второй, когда новое поколение появляется летом, приобретает бактерию от зараженных растений и передает ее здоровым. Поскольку цитрусовые не являются хозяином для бактериальных штаммов, поражающих виноград, предполагается, что распространение патогена во втором цикле является причиной массовых эпидемий болезни Пирса. Хотя экологические факторы, влияющие на возникновение эпидемий болезни Пирса в Калифорнии, еще не полностью выяснены, считается, что высокая плотность популяции переносчиков является важным компонентом этой системы. По сравнению с другими цикадками, H. vitripennis является плохим переносчиком X. fastidiosa на винограде. Возникновение эпидемий в Калифорнии в основном связано с инвазивным переносчиком, который, хотя и передает бактерию с низкой эффективностью, компенсирует это за счет очень высокой численности в близлежащих цитрусовых садах. Экологические и поведенческие характеристики H. vitripennis способствуют массовому распространению патогена на виноградниках.



