
Aperçu général du Sclerotinia sclerotiorum
Author(s): агроном Керанка Жечева, Добруджански земеделски институт – гр. Генерал Тошево, ССА; проф. д-р Иван Киряков, Добруджански земеделски институт – гр. Генерал Тошево, ССА
Date: 11.04.2025
877
![MultipartFile resource [file_data]](/assets/img/articles/заглавна-обзор-керанка.jpg)
Résumé
Sclerotinia sclerotiorum est un champignon phytopathogène qui attaque plus de 400 espèces végétales appartenant à 75 familles botaniques. Les pertes de rendement causées par cet agent pathogène peuvent atteindre 100 %. Dans les conditions de la Bulgarie, S. sclerotiorum est un ravageur clé pour un certain nombre de cultures industrielles, maraîchères et de légumineuses à graines. La présente publication fournit des informations succinctes concernant la distribution, la symptomatologie, la pathogénie et les mesures de lutte contre ce champignon.
Sclerotinia sclerotiorum (Lib.) de Bary est un agent pathogène polyphage qui attaque plus de 400 espèces végétales, principalement des dicotylédones, appartenant à 75 familles botaniques (Boland et Hall 1994). Le champignon appartient au phylum des Ascomycota, classe des Leotiomycetes. L'agent pathogène a été signalé dans plus de 100 pays en Europe, Afrique, Asie, Amérique du Nord, Amérique centrale et Caraïbes, Australie et Nouvelle-Zélande (Saharan et Mehta, 2008 ; Cohen, 2023). En Bulgarie, l'agent pathogène est un ravageur clé pour un nombre considérable de cultures industrielles, de légumineuses et maraîchères. Les dégâts causés par le champignon sont liés à l'espèce végétale, à la résistance des génotypes, aux organes attaqués ainsi qu'aux conditions édapho-climatiques, et varient considérablement, pouvant atteindre 100 % (Vasconcellos et al., 2017 ; Rather et al., 2022).

Figure 1. Symptômes causés par Sclerotinia sclerotiorum sur le haricot commun
Les maladies causées par Sclerotinia sclerotiorum portent des noms différents selon l'hôte et les organes végétaux attaqués (flétrissement sclérotinien, pourriture sclérotinienne, pourriture blanche, moisissure blanche, pourriture racinaire, pourriture de la tige) (Steadman, 1983 ; Bolton et al., 2006 ; Saharan et Mehta, 2008). Les symptômes de la maladie sont facilement reconnaissables, en raison de la formation d'un mycélium blanc cotonneux à la surface des tissus infectés (Hossain et al. 2023). Initialement, des taches imbibées d'eau de taille et de forme variables se forment sur les tissus affectés, qui se décolorent par la suite, et les tissus infectés meurent.

Figure 1a. Symptômes causés par Sclerotinia sclerotiorum sur le tournesol (forme sur tige)
Dans des conditions humides, un mycélium blanc semblable à du coton s'accumule sur les tissus affectés, qui se compacte ensuite et forme des structures noires appelées sclérotes (Fig. 1 et 1a). Des sclérotes peuvent également se former à l'intérieur des organes infectés.
Les sclérotes sont la principale source d'infection primaire. La durée de leur survie est influencée par des facteurs tels que le type de sol, l'humidité et la température, ainsi que leur position dans le sol. Il a été établi qu'en conditions sèches, les sclérotes peuvent rester viables pendant une période de 7 à 10 ans (Adams et Ayers, 1979).

Figure 2. Formation de nouveaux sclérotes (flèches) sur papier filtre humidifié.
Une expérience que nous avons menée montre que placer des sclérotes sur du papier filtre humidifié à 4°C pendant 40 jours conduit à leur développement mycélien et à la formation de nouveaux sclérotes (Fig. 2) (Zhecheva et al. 2024). Ces résultats indiquent que le champignon peut augmenter sa population en l'absence d'hôtes.
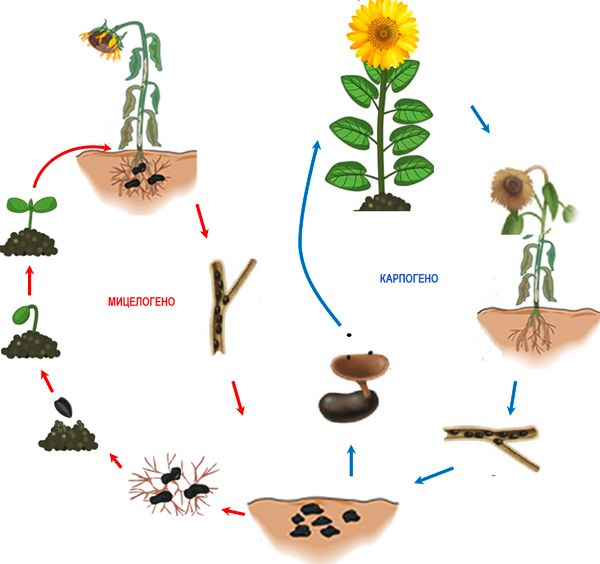
Figure 3. Cycle de vie de Sclerotinia sclerotiorum sur le tournesol
L'infection des hôtes suit deux scénarios principaux (Fig. 3). Les sclérotes situés à proximité du système racinaire ou des organes végétaux en contact avec le sol germent par du mycélium (développement mycéliogène), qui colonise superficiellement les tissus tout en produisant de l'acide oxalique (Hegedus et Rimmer, 2005 ; Hossain et al., 2023). L'acide oxalique produit supprime les mécanismes de défense des cellules tout en augmentant l'efficacité des enzymes dégradant la paroi cellulaire (CWDEs) (Hegedus et Rimmer, 2005). Le second scénario est lié à l'entrée du champignon dans un cycle sexué (développement carpogène), qui conduit à la formation de fructifications appelées apothécies (Fig. 4), à partir desquelles une quantité importante d'ascospores est libérée (Hegedus et Rimmer, 2005). Chaque apothécie peut libérer jusqu'à 10 millions d'ascospores en 7 jours, qui sont transportées par les courants d'air sur des distances de 3 à 4 km. Après s'être déposées sur les fleurs des plantes, les ascospores germent et colonisent les organes sénescents (pétales, sépales, pollen, etc.), puis attaquent les tissus adjacents. L'infection est favorisée par une humidification des plantes pendant 16 à 48 h et une température de 12 à 24 °C. La plupart des études montrent que pour que les sclérotes entrent en développement carpogène, un conditionnement préalable de plusieurs mois est nécessaire, durant lequel les sclérotes restent à une température de 0 à 5°C et à une humidité élevée (Sanogo et Puppala, 2007). La présence de précipitations et une température optimale comprise entre 20 et 25°C favorisent la formation d'apothécies et d'ascospores, mais des apothécies peuvent également se former à 5°C (Wu et al., 2008 ; Phillips, 198 ; Sanogo et Puppala, 2007 ; Godoy et al., 2017) ou à 10–15°C (Gupta et Singh, 2017). Seuls les sclérotes situés à la surface du sol ou à une profondeur de 3–5 cm forment des apothécies (Godoy et al. 2017).

Figure 4. Initiation et apothécies formées chez Sclerotinia sclerotiorum
Le nombre d'apothécies formées sur un seul sclérote dépend de sa taille et varie de quelques-unes à plusieurs dizaines. Une expérience que nous avons menée en conditions de plein champ montre que des sclérotes placés à la surface du sol et recouverts de résidus végétaux de blé et de maïs en octobre initient la formation d'apothécies fin mars (données non publiées). Aucune initiation n'a été détectée dans les variantes sans résidus végétaux et avec des résidus végétaux de tournesol. Ces résultats indiquent que la présence de résidus végétaux sur le sol favorise le développement carpogène des sclérotes en maintenant une humidité élevée.
Dans les conditions de la Bulgarie, Sclerotinia sclerotiorum présente principalement un développement mycéliogène (Saharan et Mehta, 2008 ; Genchev et Kiryakov 2002). Ce fait détermine la stratégie de lutte contre l'agent pathogène. La rotation des cultures est une mesure préventive majeure pour lutter contre le champignon (Saharan et Mehta, 2008). Il est recommandé que dans les champs où le développement de l'agent pathogène est avéré, les cultures hôtes ne soient pas cultivées pendant une période de 4 à 5 ans. Le respect des dates de semis et des densités de semis peut également limiter la manifestation et le développement de l'agent pathogène. Des semis plus précoces des cultures de printemps créent des conditions favorables à la mort des plantules et des jeunes plants à la suite du développement mycéliogène des sclérotes (O’Sullivan et al., 2021). Des densités de semis élevées créent des conditions pour une rétention prolongée de l'humidité, ce qui favorise la formation d'apothécies en cas de développement carpogène des sclérotes (McDonald et al. 2013).
L'utilisation de variétés ou d'hybrides résistants est considérée comme la mesure la plus efficace et efficiente pour lutter contre la maladie (Schwartz et Singh, 2013). La résistance à Sclerotinia sclerotiorum est de nature quantitative, ce qui complique la sélection pour la résistance (Genchev et Kiryakov, 2002 ; Schwartzand et Singh, 2013). Par exemple, la résistance des hybrides de tournesol est associée à la forme de développement de la maladie – pourriture basale, pourriture de la tige et pourriture du capitule (Castaño et al., 1993 ; Van Becelaere et Miller, 2004 ; Davar et al., 2010). Nos études montrent que les hybrides possédant une résistance à la forme sur tige sont sensibles à la forme basale de la maladie. Chez le haricot commun, deux mécanismes de résistance sont rapportés (Miklas et al., 2012 ; Schwartz et Singh, 2013). Le premier est lié au port de la plante. Les variétés à port dressé empêchent l'infection pendant la floraison grâce à une meilleure aération de la culture et à une humidité réduite. En même temps, elles positionnent les organes aériens (tiges, feuilles et gousses) à une hauteur qui empêche leur contact avec la surface du sol, ce qui prévient l'infection en cas de développement mycélien des sclérotes. Le second mécanisme de résistance est lié à des caractéristiques anatomiques des plantes qui entravent la pénétration de l'agent pathogène dans les tissus. Cette résistance est connue sous le nom de résistance physiologique (Griffiths, 2009 ; Miklas et al., 1999 ; Mkwaila et al., 2011 ; Pascual et al., 2010).
L'utilisation de fongicides est l'une des approches les plus privilégiées pour lutter contre les maladies chez les plantes cultivées. En ce qui concerne Sclerotinia sclerotiorum, l'application de fongicides est justifiée lorsqu'il existe un risque de développement carpogène de l'agent pathogène, c'est-à-dire lorsque les plantes sont attaquées pendant la floraison (Peltier et al., 2012 ; Derbyshire et Denton-Giles, 2016 ; O’Sullivan et al., 2021). Le traitement des semences avec des fongicides prévient l'infection aux premiers stades de développement des plantes (Peltier et al., 2012). En cas d'attaque directe sur la biomasse aérienne des plantes à la suite d'un développement mycéliogène des sclérotes, l'application de fongicides n'est pas efficace.
Références
1. Adams, P. B., & Ayers, W. A. (1979). Ecology of Sclerotinia species. Phytopathology, 69(8), 896-899.
2. Boland, G. J., & Hall, R. (1994). Index of plant hosts of Sclerotinia sclerotiorum. Canadian Journal of Plant Pathology, 16(2), 93-108.
3. Bolton, M.D, Thohmma, B.P.H.J. & Nelson, B.D. (2006). Sclerotinia sclerotiorum (Lib.) de Bary: biology and molecular traits of a cosmopolitan pathogen. Molecular Plant Pathology, 7(1), 1–16



